
Uomo
Uomo
(XXXIV, p. 748; App. V, v, p. 661; v. anche paleoantropologia, XXVI, p. 1; App. II, ii, p. 486; III, ii, p. 348; IV, ii, p. 720)
L'evoluzione umana
Il fattore che ha maggiormente caratterizzato l'evoluzione umana è la migrazione. I movimenti di individui, di gruppi e (raramente) di intere popolazioni sono generalmente causati da un'innovazione biologica o culturale che in un primo tempo favorisce l'aumento della popolazione nel luogo dove si verifica e, in un secondo tempo, quando il carico demografico eccede la capacità portante del territorio, provoca uno spostamento graduale, generalmente lento, di una parte della popolazione alla ricerca di nuovi territori. Questo meccanismo prende il nome di espansione demica e si distingue dalla semplice migrazione, che di per sé non implica un processo di crescita. Da un esame particolareggiato della struttura genetica di tutte le popolazioni di cui siano disponibili dati di frequenze geniche (Cavalli-Sforza, Menozzi, Piazza 1993) si è giunti alla convinzione che l'evoluzione umana, almeno nell'aspetto che si rivela attraverso i geni, è stata 'intermittente', caratterizzata cioè da una serie di espansioni demiche la cui causa è di natura culturale: per lo più la ricerca di nuove tecnologie per disporre di maggior quantità di cibo o di mezzi di trasporto più efficienti. L'effetto di queste espansioni demiche è genetico, nel senso che ancora oggi si possono ricostruire le tracce di quelle onde di spostamento nella distribuzione non casuale dei geni portati dai discendenti delle popolazioni coinvolte nelle espansioni. La diversità biologica della nostra specie si esprime quindi in una geografia di geni che riflette processi prevalentemente culturali di origine preistorica e storica.
Le espansioni demiche che hanno potuto maggiormente influire sulla nostra diversità genetica sono quelle avvenute in epoca preistorica, soprattutto durante il Paleolitico e il Neolitico, epoche in cui la densità di popolazione era così ridotta da rendere prevalente l'effetto della deriva genetica su tutte le altre pressioni evolutive. La geografia genetica generata dalla deriva genetica è una distribuzione delle frequenze geniche 'a mosaico', nel senso che regioni geografiche contigue potrebbero presentare discontinuità anche estreme. L'effetto dell'espansione demica è quello di smussare le differenze nelle aree geografiche interessate all'espansione e di generare gradienti di variazione continui come quelli che ancora oggi si osservano. Nella tabella vengono riportate le espansioni più importanti, e le innovazioni tecnologiche corrispondenti.
Espansioni nel Paleolitico
Le fonti archeologiche indicano che ci sono state almeno due espansioni di grande importanza per la storia del genere Homo. In un periodo compreso tra 1,7 e un milione di anni fa, Homo erectus ha iniziato a diffondersi dall'Africa, dove ha avuto origine, nelle aree temperate dell'Asia e dell'Europa (Klein 1989).
Homo sapiens, nella sua forma anatomica moderna (Homo sapiens sapiens), ha iniziato la sua espansione tra 100.000 e 60.000 anni fa. La scoperta in Medio Oriente di resti fossili di uomini anatomicamente moderni datati a più di 90.000 anni fa (Valladas, Reys, Joron et al. 1988) può forse cambiare l'attribuzione africana dell'origine anche di questa seconda espansione, ma Homo sapiens arcaico, morfologicamente più simile agli uomini moderni, sembra provenire dall'Africa. Accanto a questa 'ipotesi monocentrica' di entrambe le espansioni del genere Homo (Stringer, Gamble 1993), alcuni paleoantropologi (Wolpoff 1989) hanno preferito sostenere una 'ipotesi policentrica' della seconda espansione: l'u. anatomicamente moderno sarebbe il risultato di evoluzioni locali svoltesi parallelamente in Asia, Europa e Australia e iniziate con la prima espansione africana, monocentrica, di Homo erectus. I sostenitori di questa tesi adducono quale prova della sua validità la constatazione di una certa continuità morfologica dei reperti dell'Asia orientale per un arco di tempo lungo circa un milione di anni.
Le analisi dei polimorfismi genetici, sia dei prodotti del DNA (gruppi sanguigni, immunoglobuline, proteine ed enzimi) sia direttamente del DNA nucleare e mitocondriale, concordano nel separare le popolazioni africane attuali da tutte le altre, come se le prime fossero le più antiche (Cavalli-Sforza, Menozzi, Piazza 1994). Il confronto di sequenze di DNA mitocondriale in popolazioni attuali non permette una ricostruzione filogenetica sufficientemente attendibile dal punto di vista statistico (Gibbons 1992); tuttavia, pur lasciando in dubbio il luogo di origine dell'u. anatomicamente moderno (Africa o Asia Minore, appunto), tale confronto ha contribuito in modo determinante ad accreditare l'ipotesi di un'origine unica avvenuta in un'area geografica piuttosto circoscritta (Stoneking 1996). È infatti difficile spiegare altrimenti l'osservazione che i tipi di sequenze di DNA mitocondriale presentano un numero di mutazioni molto maggiore negli individui di origine africana rispetto agli individui di altre origini, come in effetti ci si attenderebbe se i primi avessero avuto antenati molto più antichi e quindi maggior tempo per differenziarsi. Sulla base dei dati genetici attuali (Cavalli-Sforza, Menozzi, Piazza 1994), nucleari e mitocondriali, non si può comunque escludere che i primi nuclei di u. moderni arrivati in Asia centrale e orientale dall'Africa o dal Medio Oriente si siano poi in parte mescolati con i discendenti locali di Homo sapiens arcaico; ciò si accorderebbe sia con l'ipotesi monocentrica sia con la continuità delle forme fossili osservata in quelle regioni.
Le cause di queste due importanti espansioni del genere Homo dall'Africa devono essere state legate a fattori di cambiamento culturale e biologico probabilmente associati sia a una ricerca sia a una manipolazione del cibo più efficienti e a una maggiore mobilità. La prima espansione dall'Africa è quella di u. con una dimensione del cervello superiore a quella dei loro predecessori e dotati di una serie di utensili in pietra relativamente più avanzati. L'evoluzione delle industrie litiche e l'uso controllato del fuoco, di cui si hanno testimonianze archeologiche che risalgono a circa 500.000 anni fa, riflettono cambiamenti culturali verosimilmente vantaggiosi in termini di disponibilità di cibo, e perciò di sopravvivenza e opportunità di espandersi in nuovi ambienti.
È probabile che una coevoluzione culturale e biologica del cervello possa aiutare a spiegare l'origine dell'u. anatomicamente moderno (Homo sapiens sapiens) e la sua successiva espansione in tutti i continenti. Il cervello umano, misurato dal volume endocranico, aveva già raggiunto le dimensioni attuali nell'u. anatomicamente arcaico (Homo sapiens; Klein 1989) prima della seconda espansione dall'Africa: durante questo periodo è possibile che il linguaggio si sia evoluto fino a raggiungere il grado attuale di precisione e complessità, e possa aver costituito il vantaggio culturale di maggior rilievo dell'u. anatomicamente moderno, capace di indurne un'espansione così estesa, nonché la sostituzione dell'u. anatomicamente arcaico, anche se in tempi diversi da luogo a luogo. Sebbene questa sia per il momento un'ipotesi di lavoro, la documentazione archeologica testimonia una notevole evoluzione dei manufatti tra 100.000 e 60.000 anni fa con un altrettanto notevole differenziamento locale delle varie industrie litiche, come se differenze di comportamento e di comunicazione avessero favorito un processo prima di segregazione, poi di divergenza di tecnologie locali in rapida evoluzione (Isaac 1976).
Un progresso tecnologico innovativo che ha favorito la diffusione dell'u. moderno è stato l'arte della navigazione. L'impiego di zattere e imbarcazioni doveva essere conosciuto almeno 60.000÷55.000 anni fa, quando l'u. anatomicamente moderno ha raggiunto l'Australia e la Nuova Guinea (Roberts, Jones, Smith 1990). Si può affermare che circa 40.000 anni fa tutto il vecchio mondo era colonizzato dalla nostra specie e che i cambiamenti culturali, di comportamento e artistici indicati dalle testimonianze archeologiche di quell'epoca testimoniano un grande mutamento. Sembra evidente che in un processo di espansione verso nuovi territori e di insediamento in nuove nicchie, un sistema preciso e veloce di comunicazione verbale debba aver conferito vantaggi non indifferenti per la sopravvivenza, favorendo così anche sotto il profilo biologico gli u. anatomicamente moderni rispetto a quelli arcaici (Cavalli-Sforza, Menozzi, Piazza 1993).
Espansioni neolitiche e postneolitiche
Sebbene alla fine del Paleolitico il mondo antico fosse già interamente abitato dalla nostra specie, gli insediamenti erano ancora di dimensioni piuttosto ridotte, con stime della popolazione totale che vanno da uno a 10 milioni di individui (Biraben 1980). Si è dimostrato con simulazioni (Rendine, Piazza, Cavalli-Sforza 1986) che i movimenti migratori avvenuti dopo il Paleolitico non hanno avuto il tempo di annullare le differenze genetiche prodotte in precedenza, ma hanno aggiunto nuovi segni ancora oggi riconoscibili nella composizione genetica delle popolazioni. Per valutare le impronte lasciate da quei fattori, rispetto alle differenze geniche originali, e ricostruire così i movimenti migratori successivi al Paleolitico occorre, tuttavia, utilizzare le informazioni combinate di un numero sufficiente di geni, perché la storia di un solo gene può descrivere eventi diversi da quelli migratori. Sono state così introdotte le mappe sintetiche (v. uomo: Razze umane, in App. V), carte geografiche in grado di riassumere in una sola immagine la distribuzione geografica delle frequenze di molti geni, al fine di riscoprire tracce di espansioni demiche che i soli documenti archeologici o linguistici non mettono chiaramente in evidenza.
Tecnicamente le mappe sintetiche sono rappresentazioni geografiche delle linee congiungenti punti, con eguale valore, delle componenti principali (in senso statistico) delle frequenze geniche rilevate in popolazioni comprese nell'area geografica considerata. La costruzione delle mappe sintetiche si basa sul metodo statistico di analisi delle componenti principali, che consente di riassumere la maggior parte della varianza statistica di un sistema multivariato in un minor numero di variabili, dette componenti principali. L'importanza di ciascuna componente è misurata dalla frazione di varianza totale a essa attribuibile: è massima per la prima componente e decresce con il rango. Le mappe sintetiche riproducono la distribuzione geografica di indici riassuntivi basate sulle medie ponderate di tutte le frequenze geniche considerate: perciò non rappresentano i dati originali di frequenza degli alleli genici, bensì l'andamento spaziale di questi indici. Le popolazioni vengono quindi rappresentate spazialmente attraverso poche nuove variabili (tre o quattro, nelle figure successivamente considerate) che concentrano un gran numero di dati originari (nel nostro caso, frequenze relative anche a un centinaio di geni).
Le mappe sintetiche della prima, seconda, terza e successive componenti principali sono cartografie riassuntive, tra loro indipendenti, della distribuzione dei geni, che spiegano porzioni sempre più piccole della variabilità genetica complessiva. Nei casi più semplici l'ordine delle componenti è associato al tempo cui la rappresentazione si riferisce, nel senso che la mappa della prima componente si riferisce comunemente a eventi di differenziamento genetico più antichi di quello rappresentato dalla seconda o dalla terza e così via. In una mappa sintetica, il processo di espansione demica si 'vede' come un gradiente continuo di valori che si sviluppa sotto forma di anelli concentrici a partire da un'area che indica approssimativamente il centro geografico dell'espansione. Poiché il processo di espansione è sempre accompagnato da altri fattori evolutivi che cambiano la distribuzione geografica della frequenza dei geni, il suo riconoscimento sotto forma di gradiente in una mappa sintetica è spesso oscuro e impossibile; tuttavia, quando il gradiente è chiaramente individuabile ed è statisticamente significativo (cioè non dovuto al caso), l'interpretazione più semplice è quella che esso rifletta la struttura genetica di popolazioni responsabili di un processo di espansione demica importante.
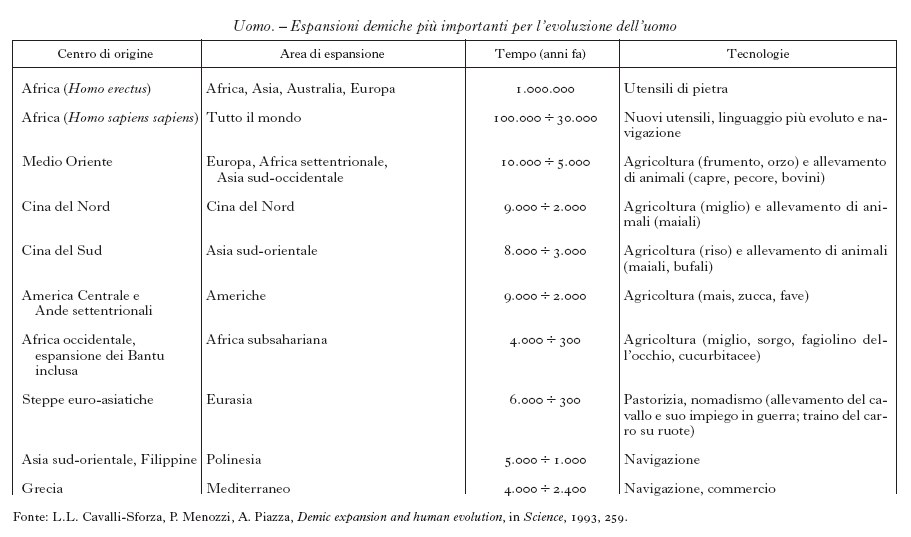
Il quadro genetico dell'Europa è riassunto nella fig. 1, che rappresenta le mappe sintetiche delle prime quattro componenti principali di 95 geni: questa cartografia recupera il 70% delle informazioni originali, nel senso che l'informazione genetica condensata nelle quattro mappe perde il 30% dell'informazione contenuta nei 95 geni considerati. La mappa della prima, e più importante, componente principale (fig. 1A) presenta un gradiente che ha origine nel Medio Oriente e si sviluppa in direzione NO. L'analogia di questa mappa con quella elaborata da A.J. Ammerman e L.L. Cavalli-Sforza (1984), in cui sono rappresentate le datazioni con ¹⁴C dei primi reperti che indicano addomesticamento di animali e coltivazione di piante in Europa, suggerisce che la diffusione dell'agricoltura nel Neolitico non è stata una semplice trasmissione culturale della tecnologia, ma ha implicato una vera e propria espansione degli agricoltori.
La seconda mappa sintetica (fig. 1B) illustra un gradiente da N verso S che potrebbe essere il risultato di un effetto del clima, ma potrebbe essere anche di origine etnica. All'estremità settentrionale vivono i Saami (Lapponi), popolazioni geneticamente simili ad altre settentrionali della Siberia, che parlano una lingua non indoeuropea ma uralica, una famiglia linguistica che oggi si trova nelle popolazioni che vivono a oriente e a occidente degli Urali (Hajdu 1962). Il gradiente genetico osservato potrebbe essere il risultato della mescolanza tra popolazioni di origine uralica e popolazioni di origine indoeuropea (Guglielmino, Piazza, Menozzi 1990), quali per es. gli Scandinavi, che si insediarono in quelle regioni dell'Europa settentrionale in epoca più recente.
La terza mappa sintetica dell'Europa (fig. 1C) suggerisce un'altra espansione con origine in una regione tra il Mar Nero e il Mar Caspio, dove l'archeologia colloca gli inizi, circa 6000 anni fa, della cultura delle tombe a kurgan. Gli allevatori nomadi delle steppe euroasiatiche avrebbero diffuso questa cultura in Europa lungo tutto il Danubio durante l'età del Bronzo (fino a circa 4500 anni fa) e gli archeologi M. Gimbutas, già nel 1970 (Gimbutas 1991), e D.W. Anthony più recentemente (Anthony, Brown 1991; Anthony 1995) l'hanno associata ai primi nuclei di popolazioni che parlavano lingue indoeuropee. Tale associazione è oggetto di controversie: l'archeologo C. Renfrew (1987) ha ipotizzato che fossero stati gli stessi agricoltori neolitici a diffondere con la nuova tecnologia anche una o più lingue della famiglia indoeuropea. L'analisi di questa mappa sintetica dei geni illustra la forza e la debolezza del documento genetico (Piazza, Rendine, Menozzi et al. 1995). Da una parte esso potrebbe risolvere il dubbio tra le due ipotesi di origine delle lingue indoeuropee: in questo caso l'espansione dell'agricoltura è documentata sia dall'archeologia sia dalla genetica. D'altra parte, poiché le rappresentazioni geografiche dei geni non sono di per sé corredate da datazioni, può sorgere il dubbio che la mappa genetica che viene attribuita all'espansione di popolazioni indoeuropee riproduca in realtà espansioni posteriori con origini geografiche simili (per es. da parte degli Sciti, che sono situati nel 500 a.C. sulle coste settentrionali del Mar Nero). Solo una ricostruzione più particolareggiata dell'archeologia delle zone interessate (in particolare la datazione di reperti che indichino l'uso del cavallo addomesticato a fini bellici e l'uso del carro dotato di ruote, probabilmente i caratteri culturali più innovativi delle prime popolazioni che parlavano lingue indoeuropee) potrebbe provare in modo definitivo la correttezza dell'ipotesi di una loro espansione a partire dall'area geografica identificata dall'analisi genetica.
La mappa della quarta componente principale (fig. 1D) dimostra un centro di diffusione in Grecia, verosimilmente attribuibile alla colonizzazione greca, che ha avuto luogo negli ultimi due millenni prima dell'era cristiana (Piazza, Cappello, Olivetti et al. 1988). Si noti come in questa mappa, a differenza delle precedenti, l'area centrale di diffusione dell'espansione corrisponda a valori più bassi (di colore meno intenso della componente principale): in effetti, quando si compongono in una sola immagine i valori di più frequenze geniche, quello che conta è il cambiamento comune e graduale delle frequenze, non il segno delle differenze che indica un andamento crescente o decrescente.
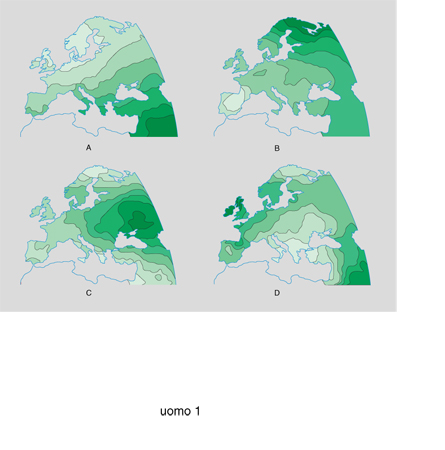
L'espansione dell'agricoltura ha avuto il suo riflesso genetico anche in Asia, nella cui prima mappa sintetica rappresentata nella fig. 2A (che riassume l'informazione di 96 geni) si riconosce un chiaro gradiente da O verso E che può interpretarsi come una semplice sintesi della progressiva diluizione del tipo genetico caucasoide nel tipo genetico orientale (o mongoloide) osservabile lungo la direzione O-E dell'Asia. Il gradiente non è regolare, ma vi è un cambiamento più brusco intorno a una linea che ha origine nella parte settentrionale degli Urali e scende verso SE per raggiungere la parte orientale dell'India: questa linea può essere considerata come il confine genetico tra tipo genetico caucasoide e tipo orientale. L.L. Cavalli-Sforza (1988) ritiene che le lingue dravidiche siano state diffuse dagli agricoltori dell'area più orientale dell'Asia Minore e siano state poi sostituite da lingue indoeuropee introdotte da popolazioni di pastori nomadi di tipo genetico caucasoide, che si sono infiltrate fino in India lasciandone intatta la parte meridionale. La mappa sintetica della seconda componente principale (fig. 2B) rappresenta un gradiente lungo la direzione N-S che forse può essere attribuito a un effetto selettivo del clima, anche se non può essere esclusa una traccia genetica delle migrazioni dal centro dell'Asia verso la Siberia. La fig. 2C illustra la terza componente della struttura genetica dell'Asia con un chiaro e interessante centro di diffusione in Giappone. Le fonti archeologiche (Koyama 1978) indicano un processo di crescita della popolazione giapponese che ha raggiunto il suo massimo (circa 300.000 individui) intorno a 4000 anni fa, durante un periodo particolarmente florido, considerando che l'economia giapponese ancora preagricola era basata essenzialmente sulla pesca. È probabile che a questo processo di crescita abbia corrisposto un'espansione (archeologicamente non documentata) di cui oggi il quadro genetico porta le tracce.
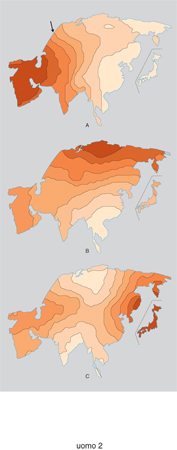
La prima mappa sintetica dell'Africa (fig. 3A), ricostruita sull'informazione di 79 geni, illustra la distinzione tra le popolazioni di origine caucasoide che abitavano a N del Sahara e le popolazioni nere che abitavano a S del Sahara. Fino a 4000 anni fa il Sahara non era un deserto e fu, anzi, teatro di importanti progressi soprattutto nelle tecniche di allevamento del bestiame. Con la desertificazione del Sahara, che può essere datata appunto a 4000÷3000 anni fa, il bestiame fu obbligato a migrare verso S, e con esso gli allevatori, nella maggior parte neri, che cominciarono a trasformarsi in coltivatori di piante, prediligendo il sorgo e il miglio, nella regione del Sahel, a sud del Sahara. Molte di queste innovazioni agricole ebbero luogo nell'Africa occidentale (Shaw 1980), da dove cominciò a espandersi poi, verso E e verso S, una corrente migratoria documentata. Questa diffusione è ben visibile nella seconda mappa sintetica (fig. 3B), dove si può osservare come la parte sud-occidentale del Sudafrica sia geneticamente diversa dall'Africa occidentale. La prima è oggi prevalentemente occupata da popolazioni che parlano lingue khoisan (Boscimani e Ottentotti) e che presentano qualche somiglianza genetica (ma non linguistica) con le popolazioni dell'Africa orientale (Nurse, Weiner, Jenkins 1985). L'area geografica che presenta il colore più chiaro definisce una regione che fu probabilmente critica per la diffusione iniziale dell'agricoltura in Africa 5000÷4000 anni fa e per l'espansione meglio conosciuta delle popolazioni agricole di lingua bantu che, originatasi nel Camerun vicino al confine con la Nigeria circa 3000 anni fa, si estese all'Africa centrale e alla maggior parte delle regioni meridionali del Sudafrica (Vansina 1984).
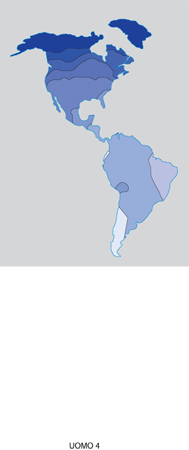
Le mappe sintetiche delle Americhe, che si riferiscono a popolazioni amerindie, sono più difficili da interpretare. Sui piccoli nuclei sopravvissuti allo sterminio dei colonizzatori europei ha agito in modo determinante la deriva genetica che deve aver generato una grande differenziazione locale. La prima mappa sintetica (fig. 4), ricostruita sull'informazione di 72 geni, illustra le differenze genetiche dei tre gruppi etnici maggiori: gli Eschimesi a N (corrispondenti alla zona con colore più scuro); i Na-dene nella zona contigua agli Eschimesi; gli Indiani d'America in tutte le altre aree con colore più chiaro. Un risultato interessante di questa tripartizione è che corrisponde esattamente a quella suggerita dall'analisi linguistica di J.H. Greenberg (1987), attualmente oggetto di accesi dibattiti tra i linguisti americani. Le altre mappe sintetiche non mostrano gradienti, ma pongono in evidenza un'estrema eterogeneità genetica: in particolare, probabilmente a causa delle vicissitudini storiche delle popolazioni amerindie, non identificano i centri ben conosciuti di diffusione dell'agricoltura del Messico e delle Ande settentrionali (Ecuador e Perù), dove il clima era più favorevole. La documentazione archeologica indica la presenza di un'economia agricola in Messico e nelle Ande a partire da circa 9000 anni fa, e una diffusione molto più tardiva, iniziata circa 4000 anni fa, verso l'America Settentrionale (Bray 1980).
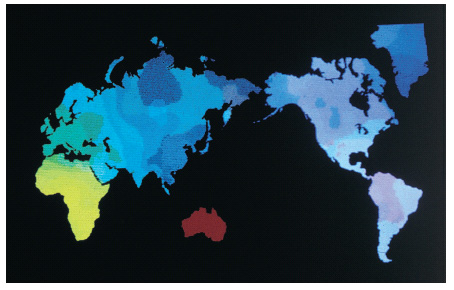
Le mappe sintetiche dei geni illustrate in precedenza si possono comporre nell'immagine geografica del mondo intero, così da fornire una fotografia genetica della nostra diversità attuale. Per rendere l'immagine ancora più informativa, le diverse componenti sono state disegnate con colori diversi e sovrapposte mediante un elaboratore di immagini. Il risultato complessivo di questa operazione di sintesi è illustrato nella fig. 5, che colloca in un'unica immagine le analisi finora effettuate. Sono chiaramente distinguibili quattro regioni etniche maggiori: Africa (giallo), Australia e Nuova Guinea (rosso), regione caucasoide (verde) e regione orientale (blu), anche se le prime due si separano dalle altre due e tra di loro in modo molto più evidente. La separazione tra Africani e Australoidi, e di entrambi i gruppi da tutte le popolazioni, concorda con l'ipotesi di un'espansione degli u. anatomicamente moderni dall'Africa. In realtà la sola distribuzione dei colori di questa mappa genetica non escluderebbe l'Australia come origine di un'espansione, ma l'evidenza archeologica dimostra che l'Africa è stata occupata molto prima dall'u. anatomicamente moderno e dai suoi antenati. Si noti poi la continuità della variazione genetica dall'Africa verso le regioni afroasiatiche caucasoidi a N del Sahara e verso il Medio Oriente; da qui verso l'Europa e verso l'Asia e dall'Asia verso le Americhe. A meno di eterogeneità locali interessanti (Cavalli-Sforza, Menozzi, Piazza 1994), la gradualità di tali variazioni indica che l'effetto delle espansioni demiche, e quindi della mescolanza di popolazioni inizialmente (ipotizziamo nel Paleolitico) molto differenziate, prevale sulle variazioni dovute a diversità ambientali. Peraltro, tale gradualità indica che la distinzione in razze geneticamente omogenee non ha alcun riscontro nei dati, un risultato che le analisi svolte e la loro traduzione in immagini sintetiche sottolineano in modo molto convincente (Piazza 1997).
Le ipotesi finora avanzate sull'evoluzione della nostra specie sono state saggiate esaminando la distribuzione geografica di frequenze geniche. Poiché i polimorfismi del DNA esaminati a tutt'oggi non eguagliano per abbondanza e dettaglio di distribuzione geografica i polimorfismi cosiddetti classici di cui ci si è prevalentemente occupati, i risultati ottenuti dai polimorfismi del DNA risultano ancora incompleti. Comunque, il quadro generale dell'evoluzione umana tracciato nelle pagine precedenti non viene smentito: le novità introdotte da questi nuovi polimorfismi arricchiscono di particolari un disegno generale sostanzialmente già delineato.
bibliografia
P. Hajdu, Finnugor népek és nyelvek, Budapest 1962 (trad. ingl. Finno-Ugrian languages and peoples, London 1975).
G.L. Isaac, Stages of cultural elaboration in the Pleistocene: possible archaelogical indicators of the development of languages capabilities, in Origins and evolution of language and speech, ed. S.P. Harnald, H.D. Staklis, J. Lancaster, New York 1976, pp. 275-88.
S. Koyama, Jomon subsistence and population, in Senri ethnological studies, 1978, 2, pp. 1-65.
J.N. Biraben, An essay concerning mankind's evolution, in Population, 1980, 4, pp. 1-13.
W. Bray, Early agriculture in the Americas, in The Cambridge encyclopedia of archaeology, ed. A. Sherrat, New York 1980, pp. 365-74.
T.S. Shaw, Agricultural origins in Africa, in The Cambridge encyclopedia of archaeology, ed. A. Sherrat, New York 1980, pp. 179-84.
A.J. Ammerman, L.L. Cavalli-Sforza, The Neolithic transition and the genetics of population in Europe, Princeton 1984 (trad. it. Torino 1987).
J. Vansina, Western Bantu expansion, in Journal of African history, 1984, 25, pp. 129-45.
G.T. Nurse, J.S. Weiner, T. Jenkins, The peoples of Southern Africa and their affinities, Oxford 1985.
S. Rendine, A. Piazza, L.L. Cavalli-Sforza, Simulation and separation by principal components of multiple demic expansions in Europe, in American naturalist, 1986, 128, pp. 681-706.
J.H. Greenberg, Language in the Americas, Stanford 1987.
C. Renfrew, Archaeology and language: the puzzle of Indo-European origins, London 1987.
L.L. Cavalli-Sforza, The Basque population and ancient migrations in Europe, in Munibe, 1988, 6, pp. 129-37.
A. Piazza, N. Cappello, E. Olivetti et al., A genetic history of Italy, in Annals of human genetics, 1988, 52, pp. 203-13.
H. Valladas, J.L. Reys, J.L. Joron et al., Thermoluminescence dating of mousterian "Proto-Cro-Magnon" remains from Israel and the origin of modern man, in Nature, 1988, 331, pp. 614-16.
R.G. Klein, The human career: human biological and cultural origins, Chicago 1989.
M.H. Wolpoff, Multiregional evolution: the fossil alternative to Eden, in The human revolution: behavioural and biological perspectives on the origins of modern humans, ed. P. Mellars, C. Stringer, Princeton 1989, pp. 62-108.
C.R. Guglielmino, A. Piazza, P. Menozzi, Uralic genes in Europe, in American journal of physical anthropology, 1990, 83, pp. 57-68.
R.G. Roberts, R. Jones, M.A. Smith, Report of thermoluminescence dates supporting arrival of people between 50 and 60 k.y. in Southern Australia, in Nature, 1990, 343, p. 133.
D.W. Anthony, D. Brown, The origin of horseback riding, in Antiquity, 1991, pp. 22-38.
M.A. Gimbutas, The civilization of the goddess, San Francisco 1991.
A. Gibbons, Mitochondrial Eve: wounded, but not dead yet, in Science, 1992, 257, pp. 873-75.
L.L. Cavalli-Sforza, P. Menozzi, A. Piazza, Demic expansion and human evolution, in Science, 1993, 259, pp. 639-46.
C. Stringer, C. Gamble, In search of the Neanderthals, London 1993.
L.L. Cavalli-Sforza, P. Menozzi, A. Piazza, The history and geography of human genes, Princeton 1994 (trad. it. Milano 1997).
D.W. Anthony, Horse, wagon and chariot: Indo-european languages and archaeology, in Antiquity, 1995, pp. 554-65.
A. Piazza, S. Rendine, P. Menozzi et al., Genetics and the origin of European languages, in Proceedings of the National Academy of Sciences of the United States of America, 1995, 92, pp. 5836-40.
M. Stoneking, Mitochondrial DNA variation and human evolution, in Human genome evolution, ed. M.S. Jackson, T. Strachan, D. Dover, Oxford 1996, pp. 263-81.
A. Piazza, Un concept sans fondement biologique, in La recherche, 1997, 302, pp. 64-68.