immunitario, sistema
immunitario, sistema
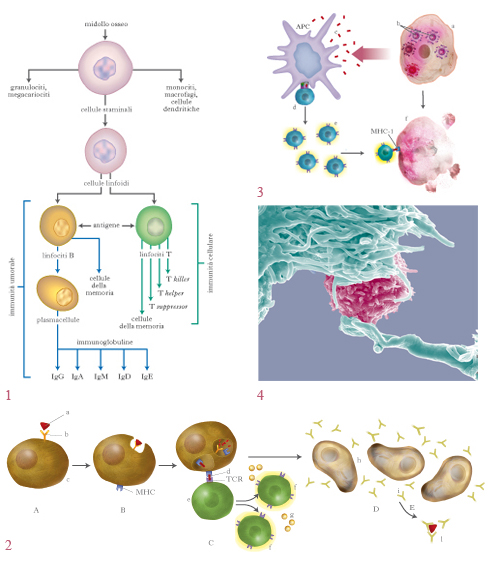
L’insieme di organi, cellule, tessuti che negli organismi superiori hanno la funzione di preservare l’organismo distruggendo o bloccando eventuali agenti esterni, quali batteri, virus, miceti, parassiti. Il sistema i. è molto complesso. Una prima distinzione al suo interno è tra le difese immunitarie (➔ immunità) innate, non specifiche (che costituiscono le prime barriere contro le infezioni) e le difese specifiche più evolute.
Filogenesi
Le risposte specifiche appaiono tardi nell’evoluzione delle specie, a livello dei primi Vertebrati, come le lamprede, pesci primitivi con bocca a ventosa. Il salto qualitativo che separa questi esseri da molluschi e artropodi avviene con la comparsa dei linfociti, cellule dotate di proprietà ‘sorprendenti’: essi sintetizzano proteine modulari (della classe delle immunoglobuline), capaci di esprimere un grandissimo numero di diversità ‘locali’ senza perdere l’omogeneità a livello di struttura molecolare portante, utilizzando tali diversità per assemblare molecole effettrici (anticorpi, recettori T) e altri recettori (detti di classe I e II) che permettono l’interazione tra cellule diverse.
Risposte umorali e cellulari
Il sistema i. specifico si divide in due branche: una realizza le risposte umorali, dirette contro antigeni extracellulari liberi (tossine, batteri, ecc.) i cui effettori sono anticorpi circolanti nei fluidi dell’organismo, e l’altra realizza risposte cellulari, i cui agenti sono linfociti capaci, tra l’altro, di riconoscere ed eliminare i virus che infettano l’organismo.
La stimolazione alla risposta
Sia la risposta umorale sia la risposta cellulare hanno inizio con la stimolazione di cellule specifiche appartenenti a un certo numero di cloni da parte di un antigene esterno (per es., una molecola proveniente dall’invasore penetrata nei fluidi dell’organismo). Nel caso della risposta umorale, il contatto tra il linfocito B specifico e l’antigene è senza intermediari. Gli antigeni non hanno una dimensione definita; solo una loro parte, chiamata epitopo, e costituita da 3 o 4 amminoacidi, viene riconosciuta e legata dal recettore specifico del linfocito B. La capacità di questo di legare l’antigene dipende dalla complementarietà spaziale, a livello molecolare, tra recettore ed epitopo, che permette l’instaurarsi di un legame non covalente di tipo intermolecolare (caratteristico delle proteine). Nelle risposte cellulari il contatto tra il linfocito T e l’antigene è mediato: l’antigene originale viene prima internalizzato da una cellula dendritica, della famiglia dei macrofagi, che processa (ossia digerisce) le molecole e ne presenta un frammento di sequenza peptidica (8÷9 unità amminoacidiche) al linfocito T, inserito nel suo recettore di classe I. Il linfocito si attiva, prolifera e genera un clone di circa duecento linfociti attivati. Se un complesso peptide-classe I uguale a quello che è servito all’attivazione del clone Tc compare sulla superficie di una qualunque cellula dell’organismo, esso denuncia la presenza di virus nella cellula.
La diversità somatica
Durante la maturazione embrionale le cellule linfatiche entrano in una fase di espansione della diversità somatica, in quanto ciascuna cellula (B e T) si costruisce il gene che codificherà il suo recettore specifico, e precisamente la parte variabile dell’anticorpo o del recettore T (esistono parti, dette costanti, uguali per tutti), ricombinando tre diversi frammenti di gene che erano già presenti nel DNA germinale. Il numero di possibili ricombinanti è enorme (milioni di miliardi), e perciò statisticamente ogni cellula è in grado di esprimere un recettore diverso da tutte le altre. La maggior parte delle specificità non sarà mai utilizzata; tuttavia, spreco e ridondanza sono il prezzo da pagare per l’efficacia della risposta immunitaria. Invece, le specificità che corrispondono agli antigeni presenti nell’organismo vengono selezionate positivamente, e l’attivazione di cloni da parte dell’antigene avviene sulla base dell’affinità, per competizione. Ciò significa che la risposta è diversa per ogni individuo, anche nel caso di gemelli uniovulari.
Il controllo
Tramite il repertorio di recettori B e T quasi infinito, il sistema i. è in grado di far fronte a qualunque agente esterno e di controbattere efficacemente alle mutazioni di quest’ultimo, in ciò favorito dalla crossreattività (➔), fenomeno per cui lo stesso recettore riconosce una rosa di epitopi simili tra loro e lo stesso antigene è riconosciuto da una rosa di recettori appartenenti a cloni simili tra loro. Una conseguenza negativa dell’allargamento delle potenziali risposte è il rischio di reagire anche – per crossreazione – contro elementi (cellule, tessuti, molecole) appartenenti all’organismo stesso (il self) generando autoimmunità (➔).
La selezione linfocitaria
La soluzione che il sistema i. ha adottato nel corso dell’evoluzione è un complesso sistema di controlli esercitati su tutti i linfociti che, generati nel midollo osseo, vengono sottoposti a una drastica selezione durante il soggiorno nel timo. La selezione timica elimina tutte le cellule i cui recettori si possono legare con alta affinità ad antigeni self. Usciti dal timo, i linfociti T helper (Th) diventano un elemento essenziale nel controllo di tutte le risposte, in partic. di quella dei linfociti B, i quali non passano per il timo. È la loro interazione con i linfociti B a permettere la moltiplicazione e la maturazione delle cellule B e la produzione di anticorpi. Invece, cellule B dirette (o crossreagenti) contro il self non trova una cellula T che interagisca con loro e perciò muoiono. L’enorme aumento in complessità del sistema i. è il costo di questo controllo di qualità della risposta, ed è in parte compensato dalla eliminazione, sempre nel timo, di un grande numero di cellule T ‘inutili’. Tutte le interazioni specifiche tra antigene, cellule dendritiche e linfociti maturi, tra linfociti T e B, tra Th e Tc (effettori citotossici), avvengono negli organi linfatici. I principali organi linfatici, oltre il timo, sono i linfonodi (collegati tra loro da una rete di vasi linfatici e la cui struttura favorisce tali incontri), la milza, le tonsille, e le placche di Peyer, inserite nella mucosa intestinale e a contatto con gli antigeni alimentari.
Risposta primaria e secondaria
Quando un dato antigene si introduce per la prima volta nell’organismo il sistema i. genera una risposta primaria (o innata), umorale, se l’antigene è extracellulare (per es., batterico) o cellulare se l’antigene è un virus. La risposta è lenta (una o due settimane), portata avanti da cloni di bassa affinità media, e poco efficace: la ragione sta nella difficoltà per le cellule effettrici (poche rappresentanti per clone) di entrare in contatto con l’antigene e superare il controllo da parte dei linfociti helper. Tuttavia, se lo stesso antigene si ripresenta (dopo settimane, mesi o anni), troverà a quel punto un organismo preparato alla risposta immunitaria, in quanto potrà contare su numerosi cloni dalle cosiddette cellule memoria, che erano state stimolate durante la risposta primaria, e soprattutto su un grande numero di cloni Th pronti a interagire con gli effettori. Il risultato è una risposta secondaria (specifica) più rapida (da qualche ora fino a 3 giorni), più ricca di cellule e anticorpi, e più selezionata ed efficiente in ragione della maturazione dell’affinità e della selezione competitiva della qualità (basata sull’affinità dei cloni).
Importanza e successi del sistema immunitario
Oltre a combattere le infezioni batteriche, virali e fungine, il sistema i. svolge un importantissimo servizio di sorveglianza che permette l’eliminazione di tessuti anomali dell’organismo e di tessuti tumorali agli stadi iniziali di crescita. Fa parte della stessa sorveglianza l’azione di rigetto dei trapianti di organi e cellule che il sistema i. considera un potenziale pericolo, fatto che costringe i trapiantati all’uso di farmaci immunodepressivi. La più importante applicazione dell’immunità specifica nella medicina preventiva è la vaccinazione, che permette di salvare migliaia di vite mediante un rinforzo mirato delle difese immunitarie.
Limiti, errori e fallimenti del sistema immunitario
Le immunodeficienze, congenite e talvolta acquisite (per es., l’AIDS), sono dovute alla malfunzione di alcuni o tutti i meccanismi di difesa. Tra gli eccessi di attività vi sono l’autoimmunità, dovuta a errori dei controlli del sistema i. o a identità casuale tra antigeni esterni e self (per es., la malattia reumatica e molte fasi autoimmuni nelle malattie del sangue e della pelle). Una reazione abnorme contro antigeni non pericolosi è alla base delle malattie allergiche (➔ allergia).