
sesso
sesso
La riproduzione sessuale è un comportamento estremamente costoso sul piano fisiologico, oltre che rischioso in termini evoluzionistici. Tuttavia, esso è ampiamente diffuso, nonostante le alternative riproduttive asessuali siano variegate e potenzialmente più efficaci. Questo paradosso rappresenta ancora oggi l’enigma della storia naturale del sesso. Il consenso scientifico ha individuato nella capacità di produrre variazione – in particolare, di accumulare mutazioni positive, di filtrare mutazioni deleterie e di difendersi dai parassiti mutando continuamente – il vantaggio selettivo principale della ricombinazione sessuale. Resta però aperto il problema di capire come il sesso sia comparso e si sia diffuso, bilanciando l’investimento richiesto. L’ipotesi più accreditata è che il successo iniziale del sesso, all’interno di piccole popolazioni, sia dovuto a una strategia variazionale di difesa in condizioni di stress ambientale e di instabilità. Un’invenzione poi rivelatasi utile in specie e in contesti selettivi diversi, al punto da condizionare profondamente la vita cognitiva ed emotiva degli animali socialmente evoluti. [➔ affetti; comportamento sessuale; cure materne, effetti dell’ambiente sulle; cure parentali; istinti; sessualità] Le due proprietà essenziali di ogni organismo sono la capacità di trasformare fonti energetiche esterne in materiale del proprio organismo (metabolismo) e di replicare le informazioni presenti nel proprio patrimonio ereditario (DNA) necessarie per ricostituirne un altro eguale. Di queste, la seconda rappresenta una proprietà che nel corso dell’evoluzione è stata caratterizzata da due opzioni di base: replicare la propria dote genetica tramite divisione in due parti dello stesso patrimonio ereditario, o ricorrere a un mescolamento dei propri geni con quelli di un partner per migliorare le possibilità di sopravvivenza. La specie umana fa parte della grande categoria di quelle che hanno scelto la seconda possibilità. Questa opzione, tuttavia, nel corso dell’evoluzione ha assistito a numerosissime varianti operative, le quali possono avere profondamente inciso a livello biologico e comportamentale sulla sessualità (➔) di ciascun individuo e anche essersi sedimentate in strati cerebrali o mentali più o meno consci (➔ istinti). Il carattere pervasivo del s. nella vita cognitiva ed emotiva degli animali sociali necessita di una comprensione evoluzionistica, al pari dei molteplici rischi ai quali esso espone gli individui, vuoi nella ricerca e nel corteggiamento del partner, vuoi nella sottrazione di tempo ed energie alla sopravvivenza, vuoi nell’esposizione alla predazione, vuoi infine nel pericolo di contrarre malattie sessualmente trasmissibili. La riproduzione per via sessuale, proprio per questo, sfida le nostre conoscenze come un rebus naturalistico: combinare la metà dei propri geni con un’altra metà proveniente da un individuo dell’altro s., per mettere al mondo una prole e moltiplicarsi, è una strategia costosa e non univoca. Come è nata e perché?
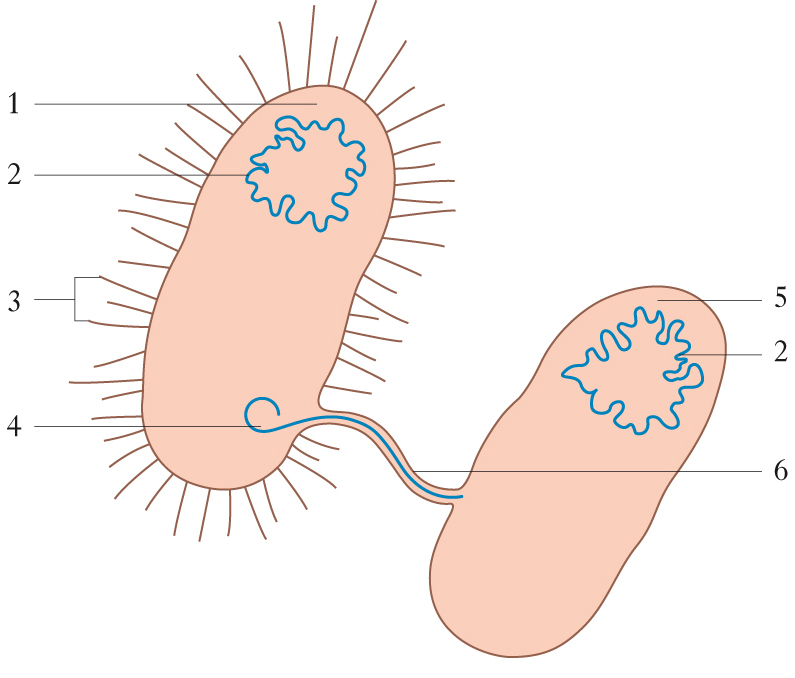
I molti modi del sesso
Numerose specie alternano il mix genetico di due sessi con la riproduzione asessuale, per es. femmine che producono uova non fecondate da cui nascono altre femmine: una discendenza di ‘amazzoni’ che fanno tutto da sole per partenogenesi. Molte piante scelgono una propagazione per via vegetativa a partire da gemme e propaggini da cui si staccano discendenti geneticamente identici. Per riprodursi un organismo può quindi clonare sé stesso, o anche più semplicemente dividersi in due, come fanno batteri e parameci. Si dà quindi in natura la possibilità di riproduzione senza s. (nei microrganismi, nelle piante, in molti invertebrati e in alcuni vertebrati), ma vale anche il viceversa, come nel ‘sesso’ dei batteri: in molti organismi unicellulari, oltre alla moltiplicazione per divisione, si assiste alla coniugazione fra due individui, cioè a uno scambio di materiale genetico in orizzontale attraverso ponti citoplasmatici. Nell’antica sessualità batterica senza riproduzione il risultato del processo è una coppia di individui geneticamente diversi da quelli di partenza. In questo modo essi acquisiscono la potenzialità di adattarsi a condizioni ambientali diverse da quelle iniziali: sembra dunque esservi un legame fra l’evoluzione del s. e la produzione di diversità.
Un’invenzione antica, ma costosa
Se però facciamo il conto di quanti esseri viventi effettivamente adottano la riproduzione sessuale (per es., la totalità di uccelli e mammiferi), o ricorrono a essa in talune circostanze, riscontriamo un indubitabile successo trasversale nell’albero della vita. Inoltre, diversamente da quanto sarebbe intuitivo immaginare, non è così sicuro che la sessualità si sia sviluppata tardivamente da una condizione primitiva di asessualità, anzi talvolta è il contrario: da scoperte recenti risulta che le specie asessuali sono spesso giovani e a volte portano con sé ancora i geni necessari per la meiosi, come se fossero evoluzioni asessuali in linee originariamente sessuali. Alcuni scienziati ipotizzano oggi che il s. abbia avuto inizio addirittura con l’antenato comune di tutti gli eucarioti. Sono state riscontrate tracce nascoste di sessualità nel DNA di organismi che si presumevano da sempre asessuali, segno che forse da tempi antichissimi vi è stata la comparsa di maschi a bassa frequenza o di forme di ricombinazione sessuale. Altre specie invece ‘prendono in prestito’ i maschi da specie affini soltanto per dare inizio allo sviluppo asessuale delle uova. finora solo i rotiferi bdelloidei, piccoli invertebrati d’acqua dolce, presentano una storia ininterrotta di partenogenesi pura che dura da quasi cento milioni di anni. Il quadro quindi si complica, perché il s. è di per sé geneticamente svantaggioso: mentre chi si riproduce asessualmente può trasmettere interamente il proprio patrimonio genetico alla discendenza, chi adotta la riproduzione sessuale può ambire solo alla metà, contenuta nei gameti aploidi dei genitori. Inoltre i maschi non contribuiscono ad aumentare il tasso riproduttivo delle femmine e il loro ruolo si limita talvolta soltanto alla fornitura di spermi per la fecondazione. Il dimezzamento secco del numero di copie di geni trasmissibili e la sopportazione del «doppio costo dei maschi», come lo definì John Maynard Smith, devono sicuramente essere controbilanciati da un qualche sostanzioso vantaggio evolutivo del s., altrimenti non si spiega la sua diffusione a partire da almeno due miliardi di anni fa.
Perché metà maschi e metà femmine
La selezione naturale può sciogliere qualche paradosso, a cominciare da quello della presenza di così tanti maschi apparentemente inutili. Se il maschio è così costoso e con la moltitudine di spermi che produce potrebbe fecondare miriadi di femmine, perché l’evoluzione non fissa una quantità minima di maschi e il resto tutte femmine? Qualche volta, in effetti, è così: nelle società cooperative di insetti eusociali – imenotteri sociali come api e formiche – le operaie femmine sono tutte sorelle e la presenza di maschi è regolata dal fatto che essi nascono solo da uova non fecondate. Nelle specie in cui fra i maschi vige una ferrea competizione per la conquista di sostanziosi harem di femmine, alle madri al di sotto di un certo peso e in condizioni non perfette, che non potrebbero allevare maschi competitivi, conviene fare figlie femmine. Poste queste eccezioni di distorsione adattativa della proporzione fra maschi e femmine, la regola in natura sembra tuttavia essere quella di una salomonica divisione a metà. Perché sopportare tutti questi maschi, quando ne basterebbe uno ogni tanto? Come notò già uno dei padri della genetica di popolazione, Ronald A. fisher, si tratta di una conseguenza della selezione naturale che agisce sul vantaggio riproduttivo del singolo individuo, anche se l’effetto complessivo non è ottimale per la specie: se in una popolazione c’è un disequilibrio a favore delle femmine, una madre in grado di generare più maschi avrà un vantaggio riproduttivo perché i suoi figli si accoppieranno più facilmente e le daranno più nipoti. Se ci fosse un gene connesso a questo tratto, avrebbe sicuro successo. Viceversa, se il numero di maschi aumenta troppo, le possibilità di accoppiarsi diminuiscono e varrà il processo inverso a favore di madri che generano più femmine. Il risultato è un bilanciamento selettivo che si stabilizza su una percentuale tendenzialmente pari fra maschi e femmine.
Le ricompense (nascoste) del sesso
Il segreto evoluzionistico del s. sembra andare oltre il piacere immediato che in alcune specie appare connesso all’atto fisico dell’accoppiamento. L’indizio fornito dal s. dei batteri è decisivo: deve esserci qualcosa di utile nel rimescolamento dei geni, nonostante la perdita di combinazioni già consolidatesi per selezione naturale e la produzione di molti genotipi intermedi. Secondo Fisher, la fusione dei genomi e la ricombinazione garantite dalla riproduzione sessuale hanno effetti sulla velocità di adattamento delle specie, perché permettono l’associazione e la diffusione delle rare mutazioni favorevoli che insorgono nelle popolazioni. In una linea asessuale, due mutazioni favorevoli per sommarsi dovrebbero comparire, con estrema improbabilità, nello stesso individuo. Se invece sono portate rispettivamente da due genitori maschio e femmina, almeno un quarto della loro prole le avrà entrambe in omozigosi. L’accumulo di riassortimenti e di combinazioni sessuali positive permette, per selezione naturale, una propagazione sufficientemente veloce delle mutazioni favorevoli. Questa funzione di ‘propellente’ per la selezione è rafforzata dal suo reciproco: la difesa da mutazioni deleterie. Il difetto maggiore della clonazione infatti è che una variazione leggermente svantaggiosa può essere risparmiata dalla selezione e trascinarsi lungo la discendenza, accumulandosi irreversibilmente insieme ad altri piccoli danni genetici. Come scoprì Alexey S. Kondrashov nel 1988, la ricombinazione sessuale permette invece di eliminare più facilmente le mutazioni deleterie perché i figli le ereditano solo in parte e la selezione naturale epura le combinazioni meno efficienti. Secondo questa teoria mutazionale, se il tasso di mutazioni leggermente deleterie in una popolazione è al di sopra di una certa soglia, il s. diventa vantaggioso e questo spiegherebbe perché risulta così difficile farne a meno da due miliardi di anni.
Il perché della dualità di generi. Una volta innescata l’evoluzione della riproduzione sessuale, si è generata nei pluricellulari una divaricazione in due generi soltanto, maschile e femminile che, a parte i funghi e pochi altri casi, è pressoché universale. La causa fondamentale di questa dualità risiede nel fenomeno dell’anisogamia, cioè del dimorfismo fra due tipologie di gameti: pochi, grandi, costosi e immobili quelli femminili; numerosi, solitamente piccoli, economici e mobili quelli maschili. Le uova rispondono alla logica evolutiva di produrre pochi gameti, ma ricchi di nutrienti che garantiscano maggiori possibilità di sopravvivenza all’embrione, mentre gli spermi alla logica evolutiva di produrre molti gameti mobili in grado di fecondare più uova possibile e di parassitarne le risorse. Le due strategie si ritrovano peraltro nella conformazione delle proli: alcune specie producono pochi figli, sui quali investire ingenti risorse parentali; altre preferiscono invece disperderne moltissimi contando sui grandi numeri. Nella riproduzione sessuale, però, le due logiche si affrontano nei due sessi della stessa specie. Da qui la radice biologica dei diversi interessi evolutivi di maschi e femmine che portano spesso a conflitti e competizioni.
L’enigma degli inizi del sesso
Lo spettro dell’inutilità imminente del maschio riaffiora, tuttavia, se pensiamo a un’altra ingegnosa soluzione riproduttiva: quella di fare tutto da soli – il maschio e la femmina – in un unico individuo che si autofeconda. Se l’incontro fisico fra uova e spermi è particolarmente difficile, e se il costo di produrre e disseminare spermi non è alto – come accade spesso per le piante e per gli animali marini fissati a un substrato –, l’ermafroditismo è un’eccellente soluzione. Alcuni pesci ermafroditi sono in grado di cambiare s. in certe fasi della loro vita. I maschi dei vertebrati terrestri sono solitamente al riparo dall’avvento dell’ermafroditismo perché la loro struttura di produzione degli spermi è troppo costosa e conviene tenere i sessi separati. Anche se le ipotesi mutazionali sono oggi suffragate da ottimi dati sperimentali, esse non risolvono un problema tipico della spiegazione evoluzionistica. La meiosi, seppur costosa, immette nelle popolazioni biologiche massicce dosi di variazione, la linfa vitale dell’evoluzione. Però, sia nel caso dell’aumento del tasso di accumulo di mutazioni favorevoli sia nel caso dell’eliminazione di mutazioni nocive, si tratta di vantaggi non immediatamente percepibili dall’individuo che si riproduce sessualmente. I loro effetti emergono nelle generazioni successive, ma sappiamo che la selezione naturale non prevede il futuro e agisce soltanto sul ‘qui e ora’ concreto della sopravvivenza differenziale. Se dunque non scoviamo un vantaggio diretto per l’individuo, ci esponiamo all’obiezione classica della ‘sovversione interna’, ossia non riusciamo a spiegare perché nel corso della storia naturale gli individui che si riproducono sessualmente non vengano puntualmente sbaragliati da quelli asessuali. Questi ultimi infatti, non avendo il costo del maschio, possono far leva su una frequenza di diffusione nella popolazione più alta. In poche decine di generazioni le femmine asessuali diventerebbero la quasi totalità della popolazione. Eppure, simili fenomeni di invasione di asessuali si sono verificati raramente in natura. Perché?
Sesso e parassiti
Se la selezione agisce subito sulla progenie, e non in previsione, la soluzione dell’enigma deve trovarsi nelle caratteristiche adattative della prole: meglio clonare individui geneticamente identici al genitore o avere discendenti diversificati fra loro? La risposta dipende dalla velocità di cambiamento dell’ambiente circostante, come suggerì William Hamilton negli anni Ottanta del secolo scorso: se il contesto delle pressioni selettive è stabile, conviene la prima soluzione; se è eterogeneo e mutevole, la seconda. Ma per compensare il costo dei maschi e offrire vantaggi immediati, l’ambiente deve cambiare a una tale velocità da essere percepibile al passaggio da una generazione all’altra e rendere così conveniente il s. già nel breve periodo. Secondo Hamilton, una componente dell’ecosistema con queste caratteristiche è la resistenza ai parassiti e alle malattie. La riproduzione sessuale garantirebbe un’incessante diversificazione genetica di generazione in generazione, obbligando gli aggressori ad aggiustare continuamente il tiro. In questo modo l’ospite, ricombinando di volta in volta il corredo genetico nei discendenti, lotta contro la sempre nutrita schiera dei suoi parassiti, che sono più veloci nel mutare e nell’aggirare le sue difese. Il dibattito resta aperto perché non è facile comparare lignaggi sessuali e asessuali che condividano ambiente e parassiti. Quando ci si riesce l’ipotesi sembra funzionare, soprattutto se i parassiti sono portati dai genitori o da parenti stretti. Ancora una volta il segreto risiederebbe nella capacità di produrre diversità, quella che manca a una popolazione uniforme di cloni, vulnerabile agli attacchi pandemici degli agenti patogeni. Il s. insomma sarebbe un buon affare in condizioni instabili di non equilibrio.
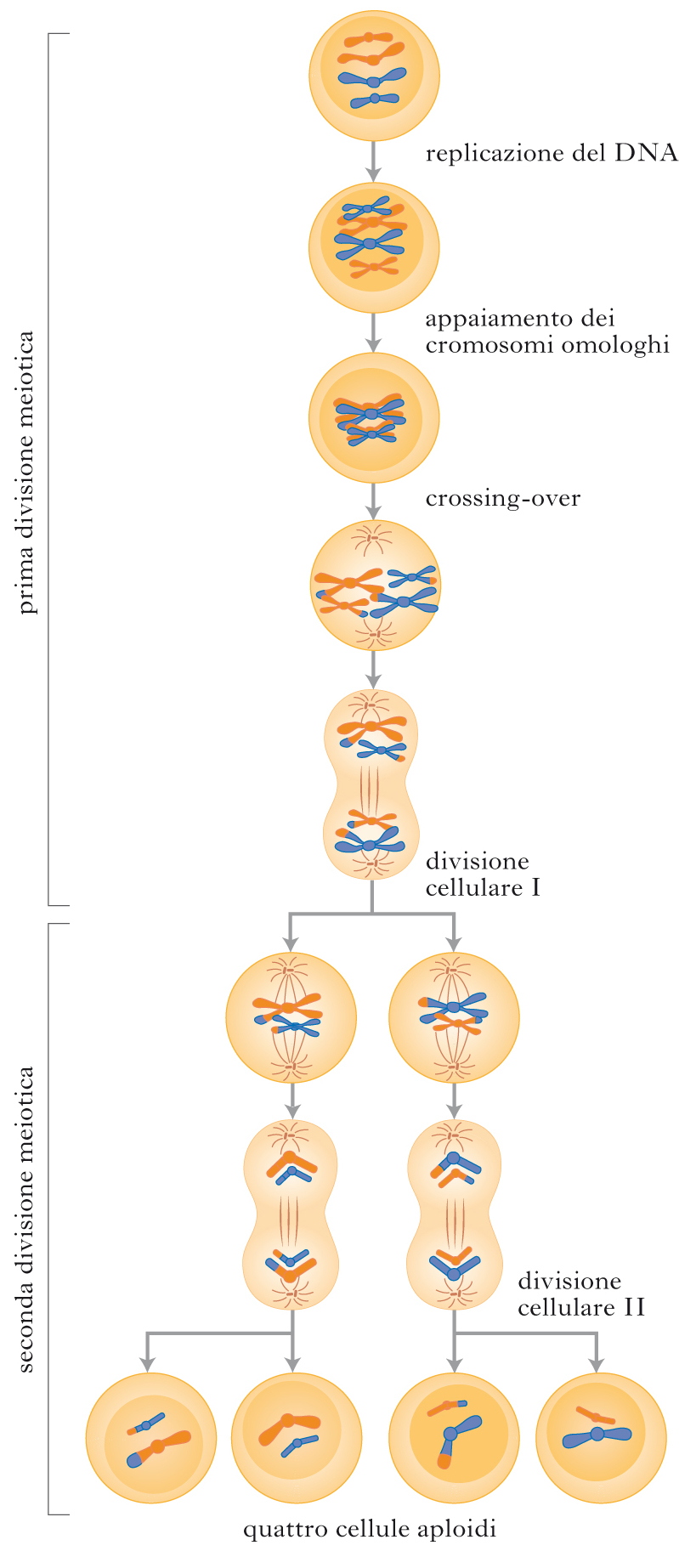
La strategia del ‘si salvi chi può’
Possiamo a questo punto immaginare un quadro esplicativo plausibile di tipo ‘pluralista’. Secondo Adam Wilkins e Robin Holliday, per capire il s. occorre comprendere l’evoluzione del sofisticato macchinario della meiosi, il quale potrebbe essere la causa innescante e non soltanto un mezzo della riproduzione sessuale. La meiosi – una volta formatesi negli eucarioti cellule diploidi con genomi sempre più grandi – sarebbe emersa come meccanismo di difesa dai rischi di ricombinazioni nocive fra cromosomi non omologhi. Da qui la fase intermedia della produzione di gameti aploidi, da unire poi più ordinatamente nella riproduzione sessuale. Anche così, però, resterebbero allettanti le soluzioni asessuali. La meiosi, tuttavia, introduce una nuova possibilità, il cui costo – secondo i modelli di Lilach Hadany e Sarah Otto – potrebbe essere stato pagato all’inizio solo occasionalmente, in specie con popolazioni frammentate ed esposte a forti stress ambientali, come estrema difesa prima di ‘abbandonare la nave’. L’indizio è dato da svariati organismi che, in effetti, optano per la riproduzione sessuale solo durante periodi di grave crisi, preferendo quella asessuale nel resto del tempo, perché il s. permette loro di produrre prole geneticamente rimescolata, in grado di adattarsi alle nuove condizioni ambientali. Gli individui capaci di girare l’interruttore genetico sulla riproduzione sessuale avrebbero dunque lasciato più discendenti. Il processo è ancor più rapido e decisivo se avviene in popolazioni di piccole dimensioni. Tutte conferme del valore variazionale del s. e del fatto che probabilmente le tre funzioni che giocano a favore della sessualità (accumulo di mutazioni positive, prevenzione da mutazioni deleterie, difesa dai parassiti) potrebbero aver agito come concause. Da qui in poi il s. ha movimentato la storia naturale, introducendo per es. disequilibri come la diffusione della promiscuità sessuale femminile e la conseguente insicurezza del maschio circa la paternità, da cui discende un’intera cascata di riadattamenti e di strategie comportamentali. Inoltre, nelle specie con una vita sociale complessa, l’accoppiamento, eterosessuale e omosessuale, non ha più soltanto una funzione strettamente riproduttiva. Le preferenze di accoppiamento fra individui dei due sessi hanno arricchito l’evolu zione con un potente meccanismo darwiniano: la selezione sessuale, forza cruciale di cambiamento capace di spiegare l’esistenza di molti caratteri appariscenti e costosi – come ornamenti e strategie di corteggiamento – che possono mettere a repentaglio la sopravvivenza fisica di chi li possiede, ma garantiscono maggiori possibilità dirette di riproduzione sessuale e quindi di trasmissione alla discendenza delle proprie caratteristiche genetiche. Charles Darwin era convinto che la reciproca scelta sessuale avesse persino plasmato gran parte delle differenze di aspetto e di comportamento degli esseri umani nei diversi popoli sulla Terra. Non mancano ancora oggi (2010) autori che assegnano alla selezione sessuale un ruolo centrale nell’evoluzione della psicologia umana. È soltanto una delle tante ricadute di un’invenzione evoluzionistica nata forse per sopravvivere a momenti di crisi ambientali acute, e rivelatasi poi sorprendentemente influente anche in contesti del tutto diversi. Telmo Pievani