
meiosi
In biologia, processo cariocinetico che determina, in alcune cellule, la riduzione dei cromosomi dal numero diploide 2n al numero aploide n, necessaria poiché, con la riproduzione sessuale e la fusione dei nuclei dei gameti, il numero dei cromosomi si raddoppierebbe a ogni generazione.
Il processo di meiosi
Negli animali la m. avviene nelle gonadi, durante la maturazione delle cellule germinali (gametogenesi): i gameti sono aploidi, tutte le altre cellule diploidi (m. gametica o terminale). Nella gran parte delle piante avviene in uno stadio intermedio del ciclo (m. sporica o intermedia), così che vi è un’alternanza di generazioni fra uno stadio diploide (sporofito o diplofito) e uno stadio aploide (gametofito o aplofito). In alcuni Protozoi e in molte piante inferiori (Clorofite, Ficomiceti, Ascomiceti, Basidiomiceti), la m. è zigotica o iniziale, cioè si verifica subito dopo la fecondazione, così che soltanto lo zigote è diploide, e tutti gli altri stadi sono rappresentati da cellule aploidi. Un nucleo con corredo cromosomico diploide contiene due copie di ciascun cromosoma, che provengono una dal padre e l’altra dalla madre tranne i due cromosomi sessuali del sesso eterogametico (per es., i cromosomi X e Y dell’uomo); i cromosomi di una coppia sono detti omologhi.
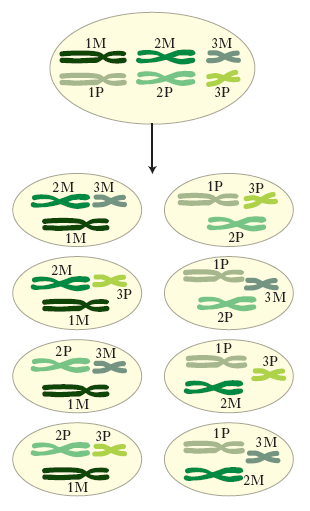
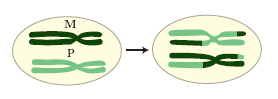
La m. è la base della variabilità genetica; si possono formare nuove combinazioni genetiche attraverso due processi fondamentali: a) i cromosomi omologhi vengono distribuiti in diverse combinazioni fra i prodotti aploidi della m. mediante un processo detto assortimento indipendente dei cromosomi. Per es., da una cellula diploide contenente tre coppie di cromosomi omologhi, 1M e 1P, 2M e 2P, 3M e 3P (dove M indica i cromosomi materni e P i cromosomi paterni), si formeranno gameti aploidi con 8 possibili combinazioni cromosomiche diverse (fig. 1). b) I cromosomi omologhi prendono parte allo scambio genetico, durante la profase meiotica, mediante il crossing-over. Un gamete aploide tipico conterrà pertanto cromosomi derivati dalla madre, cromosomi derivati dal padre e alcuni cromosomi ricombinanti che contengono informazioni derivate sia dai cromosomi paterni che da quelli materni (fig. 2). Questa cellula, con la sua specifica e unica combinazione genetica, si fonderà con una seconda cellula aploide, anch’essa con la sua specifica combinazione genetica, per produrre lo zigote diploide 2n che avrà una composizione genetica diversa da quella dei due individui diploidi da cui deriva.
Fasi della meiosi
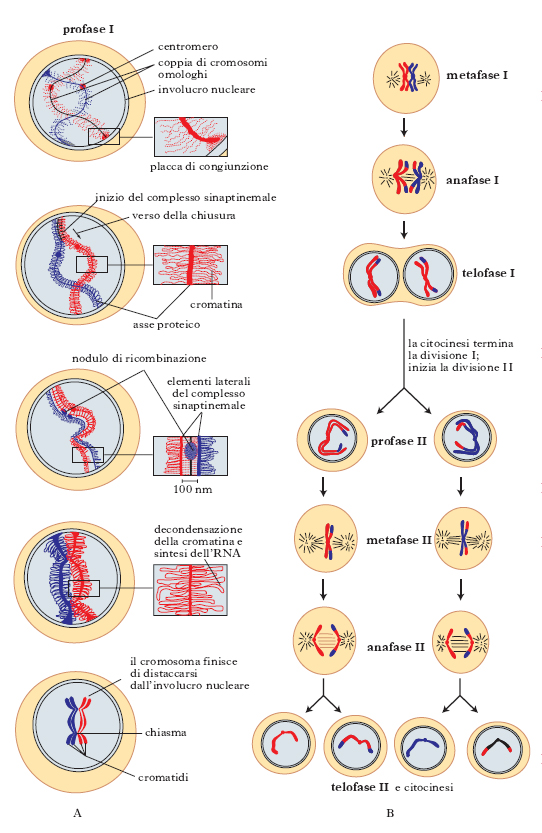
La m. è preceduta da una fase di sintesi del DNA (S); durante la m. il numero di cromosomi di una cellula diploide 2n si riduce alla metà mediante due divisioni nucleari successive: nella prima divisione i cromosomi omologhi, costituiti da due cromatidi, si separano nelle due cellule figlie, le quali ricevono un solo elemento della coppia. Nella seconda divisione meiotica si separano i due cromatidi di ciascun cromosoma per formare i 4 gameti aploidi. Ciascuna delle due divisioni è caratterizzata da 4 fasi successive: profase, metafase, anafase e telofase (fig. 3).
Profase. - La profase della prima divisione meiotica (profase I; fig. 3A) si divide in vari stadi. Nel primo (leptotene) i cromosomi si individuano al microscopio ottico e appaiono come sottili filamenti (cromonemi) in cui sono distinguibili granuli intensamente colorabili (cromomeri). Ogni cromosoma aderisce con entrambe le sue estremità all’involucro nucleare mediante una piastra di attacco. Nel secondo stadio (zigotene) i cromosomi omologhi si appaiano accostandosi per tutta la loro lunghezza con un processo a ‘chiusura lampo’ che assicura l’appaiamento di ciascun allele con il suo omologo del cromosoma opposto. La sinapsi viene preceduta dalla formazione di un asse proteico filiforme lungo ciascuno degli omologhi; durante l’appaiamento, gli assi proteici aderiscono formando una struttura a forma di scaletta, detta complesso sinaptinemale. Ciascuna coppia di cromosomi viene chiamata bivalente ma, in questo stadio, sono visibili 4 cromatidi per ciascun bivalente che pertanto può essere denominato tetrade. Segue lo stadio di pachitene, in cui i cromosomi si accorciano e si ispessiscono, sempre rimanendo appaiati. In questa fase i crossing-over determinano lo scambio fra cromatidi appartenenti ai due cromosomi omologhi diversi. Nello stadio successivo (diplotene), il complesso sinaptinemale si disgrega permettendo la separazione dei due cromosomi omologhi; essi rimangono tuttavia uniti da uno o più chiasmi, i siti dove ha avuto luogo il crossing-over. Negli ovociti, lo stadio di diplotene può durare mesi o anni perché è proprio in questo stadio che i cromosomi si despiralizzano e iniziano la sintesi dell’RNA, che servirà a fornire il materiale di riserva dell’uovo. La diacinesi è lo stadio di transizione verso la metafase quando la sintesi di RNA si arresta e i cromosomi addensati si staccano dall’involucro nucleare. Ciascun bivalente mostra i 4 cromatidi di cui è composto: i cromatidi figli di ogni cromosoma sono uniti fra loro a livello del centromero, mentre i cromatidi dei cromosomi omologhi che hanno subito il crossing-over sono legati da chiasmi.
Dopo la fine della lunga profase I, la m. prosegue con due successive divisioni nucleari (fig. 3B), che complessivamente occupano solo il 10% del tempo totale necessario per l’intero processo.
Metafase, anafase, telofase. - Nella metafase I, i bivalenti si dispongono sul fuso (sulla piastra equatoriale della cellula) e i centromeri dei due elementi di ciascun bivalente si orientano verso i poli opposti di esso. Nell’anafase I, i centromeri si allontanano verso i poli del fuso e i chiasmi si spostano verso l’estremità libera dei cromatidi, fenomeno detto terminalizzazione dei chiasmi. Nella telofase I si formano 2 nuclei con un numero aploide di cromosomi. Ciascun cromosoma è costituito dai due cromatidi uniti dal centromero. L’intercinesi, con la despiralizzazione dei cromosomi e la formazione di due cellule, è uno stadio che non sempre si verifica ed è comunque di breve durata. Segue la metafase II in cui gli n elementi di ogni nucleo, con il solo centromero ancora indiviso, si dispongono sulla piastra equatoriale. Il centromero si divide e si passa così all’anafase II, in cui i centromeri si spostano ai poli opposti del fuso e i cromatidi di ciascun cromosoma si dividono. Alla telofase II si hanno, come discendenti della cellula originaria, 4 cellule con numero aploide (n) di cromosomi. Nella gametogenesi femminile le 4 cellule prodotte sono l’uovo maturo e i 3 globuli polari, nella gametogenesi maschile sono i 4 spermatidi.
Anomalie della meiosi
La più grave alterazione durante la m. è detta non-disjunction, cioè la mancata separazione di una coppia di cromosomi (o di una coppia di cromatidi). Rilevata per la prima volta nei cromosomi X di Drosophila è legata nell’uomo a numerose sindromi: la sindrome di Down, è associata alla non disgiunzione dei cromosomi della coppia 21; la sindrome di Klinefelter e quella di Turner della coppia dei due cromosomi X; la sindrome di Pätau (trisomia 13) della coppia 13 e la sindrome di Edwards (trisomia 18) della coppia 18.