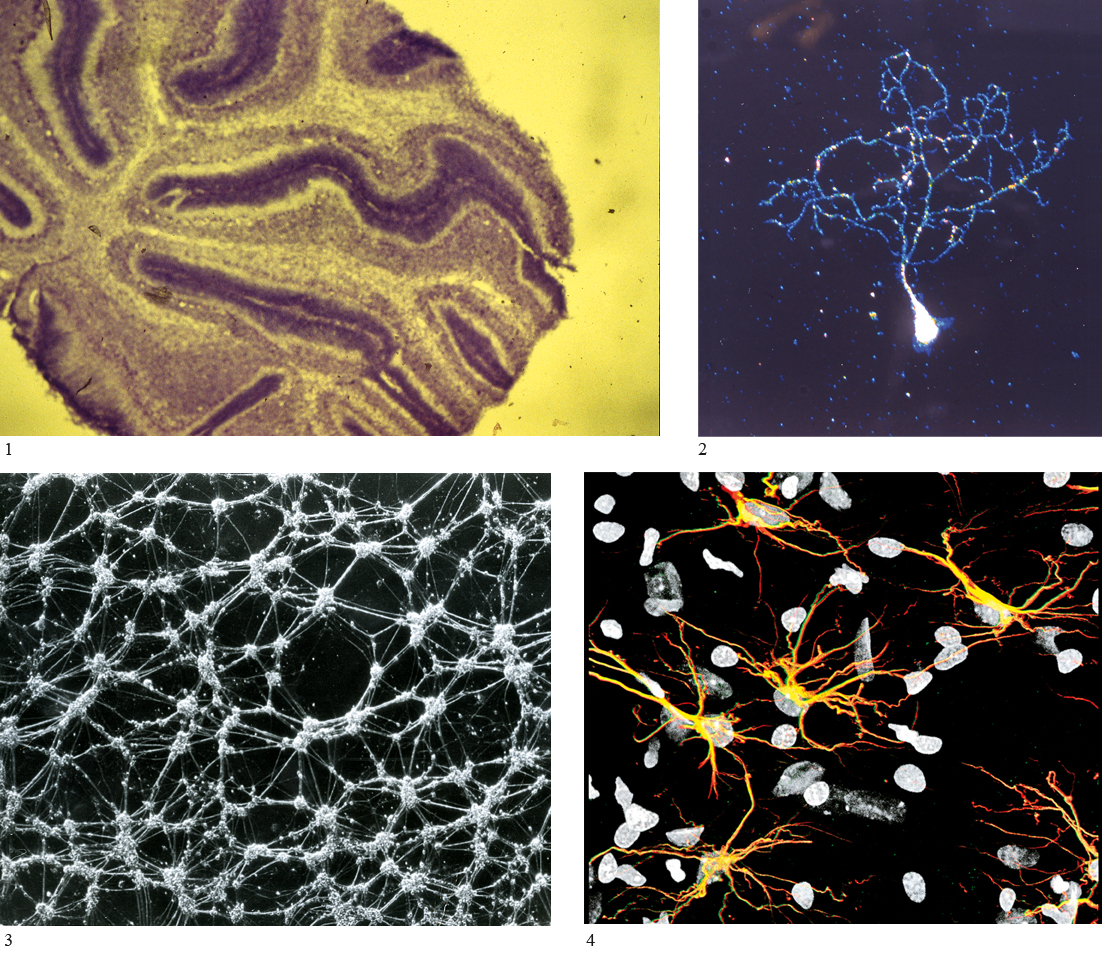
cervelletto
cervelletto
Porzione del cervello localizzata nella fossa cranica posteriore, dorsalmente al ponte del tronco encefalico. Il c. fa parte del sistema nervoso centrale ed è collegato con il tronco encefalico tramite 3 coppie di peduncoli. Il peduncolo cerebellare superiore lo collega al mesencefalo, il peduncolo cerebellare medio lo collega al ponte e il peduncolo cerebellare inferiore lo collega al bulbo. Macroscopicamente possiamo riconoscere una porzione centrale, il verme, e due emisferi cerebellari, di destra e di sinistra. Lo completano due piccole formazioni, una per lato, denominate flocculi. La superficie cerebellare ha un’organizzazione estremamente regolare e presenta una caratteristica suddivisione, a opera di fessure primarie, in lobi, che a loro volta sono suddivisi in lobuli da fessure secondarie; questi sono poi ripartiti in lamine e quindi in lamelle da fessure via via sempre più piccole, in modo da aumentare la superficie cerebellare. Si può suddividere la superficie cerebellare in 4 lobi (anteriore, medio, posteriore e flocculo-nodulare). La corteccia cerebellare è formata da sostanza grigia organizzata in moduli ripetuti. Di questi moduli fanno parte: le cellule di Purkinje, che con i loro assoni formano la via di uscita dei segnali neurali dalla corteccia cerebellare; gli interneuroni inibitori stellati, a canestro e del Golgi; infine i granuli, che costituiscono l’unico tipo di cellula eccitatoria della corteccia cerebellare e che innervano principalmente le cellule di Purkinje. Gli ingressi alla corteccia cerebellare sono costituiti dalla via rampicante, che fa sinapsi sulle cellule di Purkinje, e dalle fibre muscoidi, che sono connesse ai granuli. Entrambe queste vie sono eccitatorie. La porzione più profonda del c. è costituita da sostanza bianca, nella cui profondità si trovano tre paia di nuclei cerebellari profondi che assumono le seguenti denominazioni: nucleo del fastigio, nucleo interposito (costituito di due piccoli nuclei, nucleo globoso e nucleo emboliforme), e nucleo dentato.
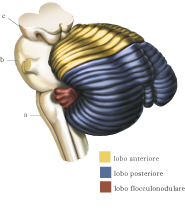
Suddivisione anatomofunzionale
È possibile suddividere il c. in tre aree diverse: l’archicerebello o vestibolocerebello, il paleocerebello o spinocerebello, e il neocerebello o cerebro cerebello. L’area dell’archicerebello è rappresentata dal nodulo (lobulo del verme) e dai due flocculi (lobuli degli emisferi) ed è l’area più ancestrale, con cui il c. è collegato, in entrambi i sensi, ai nuclei vestibolari. Questo circuito è coinvolto nel mantenimento dell’equilibrio e nel controllo dei movimenti oculari. La zona del paleocerebello è rappresentata dalla restante parte del verme e dall’area paravermina. Il verme riceve ingressi somatosensoriali, visivi, acustici e vestibolari e invia i propri segnali, attraverso il nucleo del fastigio e il nucleo interposito, ai sistemi discendenti mediali e laterali che controllano a loro volta il midollo spinale. La sua funzione principale è il controllo dell’esecuzione del movimento. La superficie restante, ossia la porzione media e laterale degli emisferi cerebellari, forma il neocerebello. Essa riceve informazioni dalla corteccia cerebrale motoria e, attraverso il nucleo dentato, invia i propri segnali alla corteccia cerebrale. È coinvolta nella pianificazione del movimento e nella formazione di memorie di procedure.
Funzione
La funzione principale del c. è di coordinare le uscite motorie: infatti, le lesioni cerebellari compromettono la coordinazione dei movimenti degli arti e degli occhi ma anche l’equilibrio. Si ritiene che il c. riceva dalla corteccia cerebrale una ‘copia’ del comando motorio che un soggetto intende volontariamente eseguire, e che riceva dagli arti informazioni relative all’effettivo svolgimento dello schema motorio impartito dalla corteccia cerebrale. Qualora sussistano differenze tra il movimento programmato e quello effettivamente realizzato, il c. è in grado di correggere, con un meccanismo di feedback negativo, il movimento durante il suo realizzarsi. Ricerche recenti hanno ipotizzato che tale compito di coordinazione del c. sia generalizzabile anche alle funzioni cognitive (per es., le lesioni cerebellari influenzano l’apprendimento di compiti di associazione di parole).
Meccanismi cellulari
Si pensa che i meccanismi di correzione della copia del comando motorio avvengano attraverso un aumento della scarica dei nuclei cerebellari. In condizioni normali, la scarica delle cellule di Purkinje inibisce i nuclei profondi. Se però non dovesse esservi correlazione tra lo schema motorio desiderato dalla corteccia e quello effettuato realmente, le fibre rampicanti, che giungono dal nucleo olivare inferiore sulle cellule di Purkinje, aumentano la loro scarica generando un forte aumento della concentrazione del calcio intracellulare nella cellula di Purkinje. Questo, sommatosi all’aumento relativo alla stimolazione delle stesse cellule di Purkinje da parte dei granuli, induce in queste ultime un processo di depressione a lungo termine (➔) con conseguente diminuzione di scarica delle cellule di Purkinje e disinibizione dell’attività dei nuclei profondi. Così il movimento tenderà a essere corretto tramite le vie di controllo che dai nuclei profondi si portano alla corteccia o, attraverso le vie motorie discendenti, al midollo spinale.