
percezione del colore
percezione del colore
Qualità della sensazione visiva costruita dal nostro cervello. La p. del c. è considerata un qualia della mente (ossia un aspetto qualitativo della vita mentale). Tuttavia il processo è strettamente legato a una particolare proprietà della luce, la lunghezza d’onda. La visione del colore nell’uomo (e nei primati catarrhini o scimmie del vecchio mondo) inizia con il processo di assorbimento della luce da parte di tre distinti fotorecettori (coni) contenenti il fotopigmento (molecola che ha la proprietà di assorbire la luce) con tre diversi spettri di assorbimento. I coni – detti L, M e S perché hanno picchi di assorbimento a lunghezze d’onda lunghe (580 nm), medie (540 nm) e corte (430 nm) – hanno spettri di assorbimento molto ampi in sovrapposizione fra loro: non esiste infatti una luce di lunghezza d’onda che riesca a eccitare solo uno dei tre coni. Come aveva intuito Hermann von Helmholtz nell’Ottocento, la nostra p. del c. nasce dal paragone delle attività relative dei tre coni, che generano la stessa risposta con un’infinità di combinazioni di spettri luminosi. Un’uguale sensazione di un colore (colori metamerici) può essere generata da un’ampia gamma di spettri, purché queste generino la stessa terna di attività dei coni L, M e S. Per es., la stessa sensazione di giallo può essere ottenuta utilizzando una luce di lunghezza d’onda corrispondente al giallo (ca. 580 nm) addizionata a una luce bianca (spettro quasi piatto), oppure dalla somma di una luce con lunghezza d’onda corrispondente al verde con una corrispondente al rosso. Questo principio di visione tricromatica (perché ottenuta dalla stimolazione di tre fotorecettori) è stato anche utilizzato nel progettare televisori e videocamere a colori.
Tinte uniche
Al di là della combinazione additiva delle lunghezze d’onda, esistono sensazioni di colori che hanno una valenza cardinale, le cosiddette tinte uniche: blu, verde, giallo e rosso. Sono uniche perché nessuno di questi colori è percepito come se contenesse una parte degli altri. Ewald Hering intuì che ciò era dovuto a un effetto di antagonismo fra meccanismi neuronali, ormai ben conosciuti, chiamati sistemi cromatici opponenti: l’adattamento cromatico svela facilmente la loro esistenza. Se si osserva per un lungo tempo un sfondo colorato, la successiva presentazione di uno sfondo bianco genererà una percezione di colore. Se lo sfondo era rosso, lo sfondo bianco apparirà di un leggero verde acquerello, se blu, giallo e così via. Questi effetti sono mediati da specifiche selettività neuronali. Per es., i segnali dei coni L e M convergono sui neuroni gangliari retinici in modo opponente: se un cono L eccita il neurone gangliare questo riceve anche un’inibizione da un gruppo di coni M. Si formano così risposte selettive per modulazioni cromatiche lungo due direzioni che sono cardinali per l’analisi della p. del c. nell’uomo, ovvero per variazioni dell’asse rosso-verde e dell’asse blu-giallo.
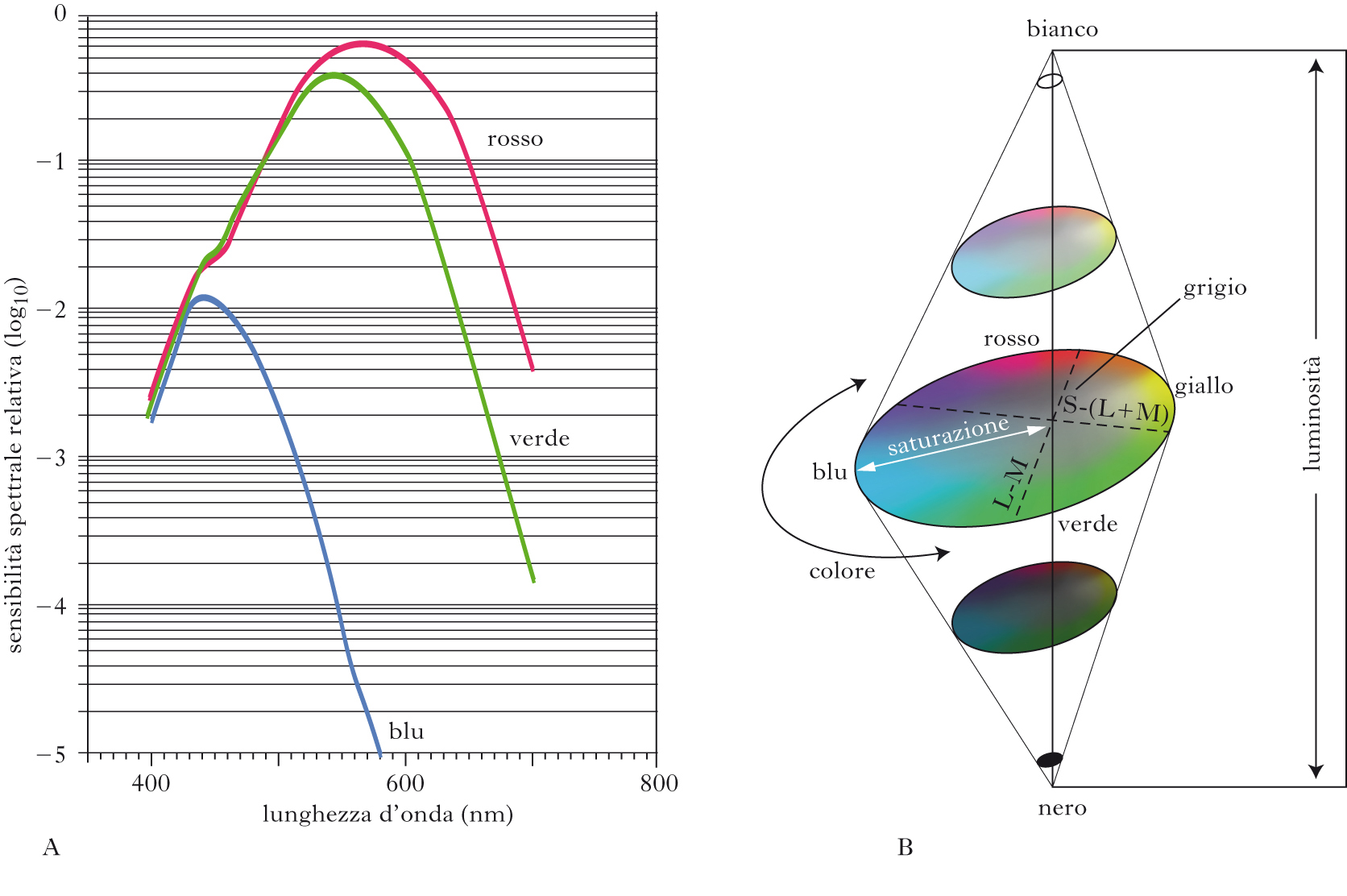
Rappresentazione della percezione cromatica
Tutti gli stimoli definiti solo da variazioni spaziali o temporali di colore (e quindi a luminanza costante ovvero equiluminanti) possono essere rappresentati su cerchi (sezioni della sfera dei colori) in cui i due assi ortogonali corrispondono alle lunghezze d’onda che eccitano solo il sistema opponente L-M (rosso-verde) e il sistema S-(L+M) (blu-giallo). Questi cerchi sono sezioni della sfera dei colori, e l’azimut delle sezioni corrisponde a variazioni di stimoli diversi per luminanza: modulazioni parallele all’asse z corrispondono a stimoli modulati solo in luminanza. I tre attributi del colore, ovvero la tinta (o tonalità di colore), la saturazione (chiamata anche purezza o croma) e la luminosità (o brillanza) sono espressi in questo spazio dalle coordinate sferiche.
Contrasti cromatici e variazioni di luminanza
I contrasti cromatici sono attualmente definiti sulla base delle attività di questi meccanismi opponenti e quindi sono specifici per i due assi cardinali, e non per i valori colorimetrici della luce. È utile puntualizzare che se si utilizzano queste definizioni funzionali di contrasto cromatico si verifica che il sistema visivo è ugualmente efficiente a rilevare la presenza di stimoli equiluminanti o modulati in luminanza, avendo anche un’acuità visiva (➔) simile per queste due classi di stimoli purché il contrasto a livello dei coni sia uguale. La visione di stimoli equiluminanti ha caratteristiche diverse rispetto a stimoli modulati in luminanza. La visione del movimento e della profondità di questi stimoli è particolarmente deficitaria, mentre i processi di segmentazione di una scena visiva in oggetti o superfici sono facilitati in presenza di informazioni cromatiche.
Alterazioni nella percezione dei colori
Gli oggetti del mondo reale assorbono energia in particolari intervalli di lunghezze d’onda, così che lo spettro della luce riflessa risulta diverso da quello della luce incidente. Lo spettro della luce riflessa può variare enormemente in funzione della luce utilizzata come illuminante, eppure normalmente la p. del c. delle superfici di un ambiente non varia. Questo fenomeno, detto della costanza dei colori, è una proprietà del sistema visivo legata alla valutazione del contrasto cromatico degli oggetti rispetto allo sfondo e richiede un’elaborazione complessa e globale della scena. Questa elaborazione sembra essere effettuata a livello corticale in aree associative specializzate. Lesioni in queste regioni corticali portano a una perdita selettiva del riconoscimento della tinta e della visione cosciente del colore. Questa patologia, detta acromatopsia corticale, è tuttavia molto rara. Alterazioni più comuni della visione del colore riguardano anomalie delle curve di assorbimento dei fotopigmenti o l’assenza di un particolare fotopigmento (daltonismo). In questi casi la discriminazione dei colori è fortemente alterata, e in caso di assenza di uno dei fotopigmenti aumenta lo spazio dei colori metamerici. Altre alterazioni sono legate ai processi nervosi che trasmettono le informazioni cromatiche (la via parvocellulare e koniocellulare) e in questo caso viene alterata la sensibilità per il contrasto cromatico, ma non necessariamente la composizione dei colori metamerici.